Аутофагия как ингибитор инициирования и развития рака
Первая заслуживающая внимания связь аутофагии с раком появилась, когда лаборатория Бет Левайн (Beth Levine) идентифицировала Beclin 1 как человеческий гомолог Atg6 дрожжей на основании его способности взаимодействовать с онкогеном BCL2. Эта работа также предполагает, что Beclin 1, который часто делетирован при опухолях человека, может функционировать как супрессор опухолей. Однако важность гена BECN1 в раке человека ставится под сомнение, поскольку у людей он находится рядом с хорошо известным супрессором опухолей BRCA1; это вело к предположению, что геномная потеря, которая часто наблюдается в опухолях человека, на самом деле является лишь следствием этой близости, а не признаком того, что сам Beclin-1 важен для супрессии опухолей. Однако инактивация одной копии гена Beclin 1 у мышей сама по себе достаточна для индукции рака, что свидетельствует о том, что, по меньшей мере, у мышей Beclin 1 может функционировать как истинный опухолевый супрессор. Проонкогенные эффекты генетической делеции других регуляторов аутофагии у мышей, включая AMBRA-1, Bif-1 и Atg4c, а также делеция Atg5 и Atg7 также вызывают формирование опухоли. Интересные загадки также возникли из этих исследований. Например, у мозаичных Atg5-/- животных опухоли (которые имели тенденцию оставаться небольшими и не прогрессировали) были обнаружены только в печени, несмотря на то, что клетки с дефицитом аутофагии встречались и в других тканях животного.
Многие факторы риска развития рака, включая старение, хроническое воспаление и ожирение, связаны со снижением аутофагии. Хотя такая связь предполагает, что дефектная аутофагия способствует увеличению риска заболевания раком, часто трудно строго продемонстрировать, что такие ассоциации являются причинными. Однако в настоящее время все согласны с тем, что, хотя нам еще многое предстоит узнать относительно основных механизмов, аутофагия может предотвратить возникновение опухоли (рис. 3). Эта способность включает в себя несколько механизмов. Активность супрессора опухоли обусловлена, по меньшей мере, частично способностью аутофагии удалять старые и поврежденные органеллы, такие как митохондрии и пероксисомы, которые могут генерировать генотоксические реактивные окислительные вещества (АФК). Аутофагия также удаляет поврежденные белки, которые в противном случае могли бы вызвать усиление стресса эндоплазматической сети (ER). Эти эффекты, в свою очередь, приводят к тому, что аутофагия способна уменьшить мутации генома и другие повреждения клеток. Аутофагия также участвует в старении опухолевых клеток, которые могут блокировать прогрессирование опухоли.
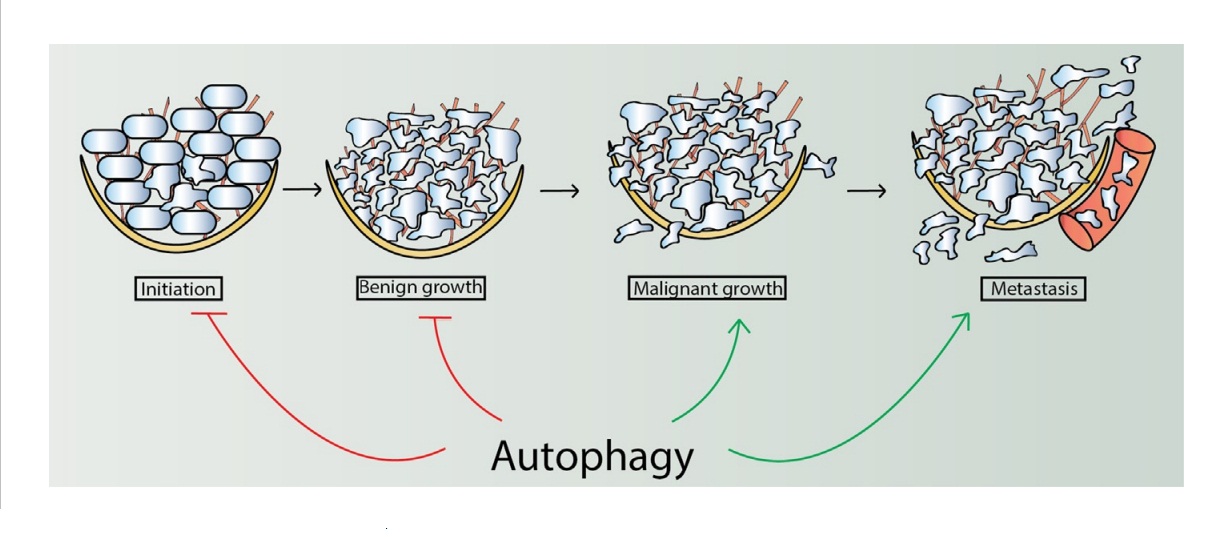
Фиг. 3. Аутофагия играет сложную роль в развитии опухолей, когда она может супрессировать опухолевый рост на ранних стадиях развития заболевания, но способствует прогрессированию опухоли на более поздних стадиях.
Способность аутофагии удалять специфический внутриклеточный материал также играет роль в ее способности защищать от рака. Хорошим примером является то, что вызывающие рак инфекционные агенты, такие как Helicobacter pylori, можно устранить с помощью селективной формы аутофагии, называемой ксенофагией. Способность аутофагии контролировать уровни p62 также влияет на возникновение рака, поскольку p62 может действовать как про-онкогенный белок.
Помимо нацеливания на специфические субстраты для аутофагии, p62 является важной сигнальной молекулой, которая регулирует NFkB и фактор транскрипции, связанный с NF-E2-фактором 2 (NRF2), который является регулятором транскрипции антиоксидантных генов. p62 также важен при хронических воспалительных состояниях, которые способствуют развитию рака, включая панкреатит и воспаление печени. Одно исследование прямо указывает на то, что p62 является критическим медиатором роли аутофагии в регуляции развития рака, поскольку развитие опухоли печени, вызванное делецией гена Atg7, снижается при одновременном удалении гена p62.
Аутофагия как промотор прогрессирования и метастазирования рака
Большинство современных данных свидетельствуют о том, что после образования опухолей первичная роль аутофагии заключается в том, чтобы способствовать прогрессированию рака. На нескольких моделях мышей было продемонстрировано, что потеря регуляторов аутофагии (часто, но не исключительно путем делеции генов Atg5 или Atg7) в установленных опухолях может снизить рост опухоли и повысить гибель раковых клеток. В опухолях легких также было показано, что делеция важных генов аутофагии может привести к переходу от более агрессивных аденокарцином к менее агрессивным типам опухолей, в частности к онкоцитомам, которые связаны с повышенным уровнем поврежденных митохондрий из-за отсутствия митофагии.
В большинстве исследований на животных, в которых изучалось влияние инактивации регуляторов аутофагии на поведение опухоли, нокаут гена аутофагии происходил одновременно с генетической мутацией, которая управляет развитием опухоли. Это затрудняет определение того, существуют ли разные роли для аутофагии на разных этапах эволюции опухоли. Эти исследования также плохо моделируют эффекты клинического вмешательства, поскольку большинство пациентов не получают лечения до тех пор, пока у них не появится установленная опухоль.
Для решения этой проблемы использовались более сложные модели на животных, где ингибирование аутофагии происходит после развития опухоли. В одном исследовании это позволило определить эффекты полного торможения аутофагии всего тела при нокауте Atg7 до или после развития рака легких, вызванного KRAS. Этот подход моделирует сценарий, в котором вводится идеальный ингибитор аутофагии, который воздействует на все клетки организма и непосредственно воздействует на механизм аутофагии. Эффекты были драматичными и ясно демонстрировали, что полное торможение аутофагии у животного является равномерно летальным, но с различной кинетикой. Некоторые мыши умерли в течение нескольких дней, в то время как другие умерли через 2-3 месяца. Первая группа умерла из-за инфекции Streptococcus, что согласуется с известной ролью ксенофагии в устранении патогенного Streptococcus из клеток. Все остальные мыши умерли в течение нескольких месяцев из-за нейродегенерации. Опять же, этого можно было ожидать, потому что аутофагия защищает от нейротоксичности, вызванной агрегированными белками. Кроме того, а также, как и следовало ожидать, мыши с делецией Atg7 были очень чувствительны к питательному стрессу, так как голодание в течение 24 часов, стресс, который мыши дикого типа могут пережить без проблем, был смертельным для большинства животных. Эта токсичность может свидетельствовать о том, что аутофагия может быть слишком токсичной, чтобы быть полезной для терапии рака. Тем не менее, когда авторы изучали эффект полного подавления аутофагии в организме после образования опухоли, особенно во время, когда животные были здоровы до нейродегенерации, они обнаружили, что опухоли легких регрессировали. Интересно, что противоопухолевый эффект был значительно выше, когда Atg7 был удален во всех тканях, по сравнению с эффектом, наблюдаемым, когда Atg7 был удален только в опухолевых клетках. Это говорит о том, что аутофагия имеет важные эффекты в неопухолевых клетках, которые влияют на поведение опухолевых клеток. Таким образом, хотя аутофагия действительно важна для выживания организма, может существовать потенциальное терапевтическое окно, в котором ингибирование аутофагии может уничтожить опухоли. Нокаутные гены, которые являются постоянными и необратимыми, не будут иметь такой же токсичности, как лекарства, которые могут быть остановлены на определенный период времени и вряд ли когда-либо полностью подавят процесс. Другая модель использовала индуцибельную экспрессию доминантно-негативной мутантной молекулы ATG4, которая блокирует аутофагию на мышиной модели рака поджелудочной железы. Эта модель важна, потому что она позволяет обратимое ингибирование аутофагии путем удаления индуктора, который активирует экспрессию доминантного негатива, а также допускает прерывистое лечение, которое лучше имитирует способ использования фармакологических ингибиторов процесса. Важные выводы из этого исследования заключались в том, что ингибирование аутофагии эффективно для ингибирования роста опухоли (то есть аутофагия необходима для прогрессирования и поддержания опухоли) и что эти эффекты включают как автономные, так и неавтономные механизмы опухолевой клетки, что поднимает вопрос о том, что именно эти механизмы находятся.
Что касается его способности подавлять возникновение опухоли, было выявлено множество механизмов, с помощью которых аутофагия может способствовать прогрессированию опухоли, и вполне вероятно, что многие различные механизмы играют роль в конечном поведении опухоли. Раковые клетки обычно чувствительны к апоптозу, и один важный механизм, с помощью которого аутофагия защищает выживание опухолевых клеток, заключается в поддержании опухолевых клеток ниже их апоптотического порога. Опухолевые клетки часто подвергаются питательному стрессу и гипоксии; Известная роль аутофагии в защите клеток от этих стрессов может способствовать выживанию опухолевых клеток в условиях стресса. Также было показано, что аутофагия играет важную роль в подвижности опухолевых клеток благодаря множеству механизмов, включая аутофагий-зависимую секрецию цитокинов, обмен белков цитоскелета, которые важны для регуляции подвижности клеток и регуляции малых GTPases семейства Rho. Сообщалось, что аутофагия важна для поддержания активности раковых стволовых клеток и для выживания спящих опухолевых клеток. Все эти механизмы могут способствовать увеличению метастазирования, и есть многочисленные сообщения о том, что у клеток метастатического рака повышена аутофагия. Большинство из этих эффектов являются автономными для опухолевых клеток, однако было проведено несколько исследований, которые показывают важные автономные функции неопухолевых клеток для аутофагии. Наиболее понятный пример показывает, что аутофагия в микроокружении опухоли (например, звездчатые клетки поджелудочной железы) продуцирует аминокислоты, которые используются для питания метаболических путей в раковых клетках.
Аутофагия и иммунный ответ на канцер
Другой важный аспект влияния аутофагии на прогрессирование опухоли связан с аутофагией и иммунным ответом рака. Однако, как и в случае гибели клеток, были выявлены как функции, стимулирующие опухоль, так и функции, подавляющие опухоль. Аутофагия может стимулировать противоопухолевые иммунные ответы через несколько механизмов. Адаптивный противоопухолевый иммунный ответ требует, чтобы антигены опухоли распознавались иммунной системой. Аутофагия играет важную роль в презентации антигена молекулами МНС класса I и II Т-клеткам. Аутофагия также важна для обеспечения созревания и выживания Т-клеток. Активация эффективного ответа Т-клеток на опухоли требует, чтобы раковые клетки умирали в результате «иммуногенного» типа смерти, который включает воздействие кальретикулина на поверхность умирающей клетки и высвобождение связанных с повреждением молекулярных структур (DAMP), таких как HMGB1. как спс. На высвобождение DAMP и ATP влияет аутофагия в умирающей клетке. Все эти эффекты предполагают, что аутофагия необходима для создания эффективного противоопухолевого иммунного ответа. И наоборот, аутофагия может работать против других форм иммунного контроля рака, например, уменьшая способность естественных киллеров (NK) убивать раковые клетки. Появляющиеся данные о неразрушающей роли аутофагосомных лесов в апоптозном / некроптотическом переключении в гибели клеток добавляют новые вопросы для рассмотрения, поскольку некроптоз особенно эффективен в индукции противоопухолевого иммунитета.